1.前期Ⅰ 这一时期发生在核内染色体复制已完成的基础上,整个时期比有丝分裂的前期所需时间要长,变化更为复杂。根据染色体形态,又被分为5个阶段:
(1)细线期(leptotene)细胞核内出现细长、线状的染色体,细胞核和核仁继续增大。在一些植物中,细长的染色体还经过一度缠绕、缩短变粗,使轮廓清晰可见。这时,每条染色体含有二条染色单体,它们仅在着丝点处相连接。
(2)偶线期(zygotene)也称合线期。细胞内的同源染色体(即来自父本和母本的二条相似形态的染色体)两两成对平列靠拢,这一现象也称联会(synapsis)。如果原来细胞中有20条染色体,这时候便配成10对。每一对含4条染色单体,构成一个单位,称四联体(tetrad)。
(3)粗线期(pachytene)染色体继续缩短变粗,同时,在四联体内,同源染色体上的一条染色单体与另一条同源染色体的染色单体彼此交叉组合,并在相同部位发生横断和片段的互换,使该二条染色单体都有了对方染色体的片段,从而导致了父母本基因的互换,但每个染色单体仍都具有完全的基因组。
(4)双线期(diplotene)发生交叉的染色单体开始分开,由于交叉常常不止发生在一个位点,因此,使染色体呈现出X、V、8、0等各种形状。
(5)终变期(diakinesis)染色体更为缩短,达到最小长度,并移向核的周围靠近核膜的位置。以后,核膜、核仁消失,最后并出现纺锤丝。
2.中期Ⅰ 各成对的同源染色体双双移向赤道面。细胞质中形成纺锤体。这时与一般有丝分裂中期的区别在于有丝分裂前期因无联会现象,所以中期染色体在赤道面上排列不成对而是单独的(图1-37)。
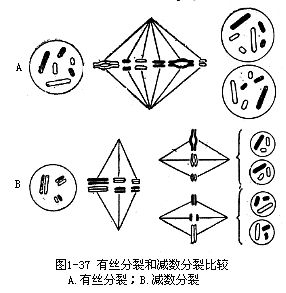
3.后期Ⅰ 由于纺锤丝的牵引,使成对的同源染色体各自发生分离,并分别向两极移动。这时,每一边的染色体数目只有原来的一半。
4.末期Ⅰ 到达两极的染色体又聚集起来,重新出现核膜、核仁,形成二个子核;同时,在赤道面形成细胞板,将母细胞分隔为两个子细胞。由上可知,这两个子细胞的染色体数目,只有母细胞的一半。然后,新生成的子细胞紧接着发生第二次分裂。也有新细胞板不立即形成,而连续进行第二次分裂的。
(二)第二次分裂(简称分裂Ⅱ)
分裂Ⅱ一般与分裂Ⅰ末期紧接,或出现短暂的间歇。这次分裂与前一次不同,在分裂前,核不再进行DNA的复制和染色体的加倍,而整个分裂过程与一般有丝分裂相同,分成前、中、后、末4个时期,前期较短,而不像分裂Ⅰ那样复杂。
1.前期Ⅱ 核内染色体呈细丝状,逐渐变粗短,至核膜、核仁消失。
2.中期Ⅱ 每个细胞染色体排列在赤道面上,纺锤体明显。
3.后期Ⅱ 每条染色体的二条染色单体随着着丝点的分裂而彼此分开,由纺锤丝牵向两极。
4.末期Ⅱ 移向两极的染色单体各组成一个子核,并各自形成一个子细胞。至此,整个减数分裂过程正式完成。
由上可见,减数分裂中一个母细胞要经历两次连续的分裂,形成4个子细胞,每个子细胞的染色体数只有母细胞的一半。染色体的减半实际上是发生在第一次分裂过程中。
减数分裂具有重要的生物学意义。减数分裂是与生物的有性生殖相联系的,它发生在特殊的细胞中,通过减数分裂导致了有性生殖细胞(配子)的染色体数目减半,而在以后发生有性生殖时,二个配子相结合,形成合子,合子的染色体重新恢复到亲本的数目。这样周而复始,使有性生殖的后代始终保持亲本固有的染色体数目和类型。因此,减数分裂是有性生殖的前提,是保持物种稳定性的基础。同时,在减数分裂过程中,由于同源染色体发生联会、交叉和片段互换,从而使同源染色体上父母本的基因发生重组,从而产生了新类型的单倍体细胞,这就是有性生殖能使子代产生变异的原因 。