第六节 种子和果实
被子植物的受精作用完成后,胚珠便发育为种子(图4-57),子房发育为果实。有些植物,花的其他部分和花以外的结构,也有随着一起发育成果实的一部分的。

种子是所有种子植物特有的器官。种子植物中的裸子植物,因为胚珠外面没有包被,所以胚珠发育成种子后是裸露的;被子植物的胚珠是包在子房内,卵细胞受精后,子房发育为果实,里面的胚珠发育成种子,所以种子也就受到果实的包被。种子有无包被,这是种子植物中裸子植物和被子植物两大类群的重要区别之一。种子植物除利用种子增殖本属种的个体数量外,同时也是种子植物借以渡过干、冷等不良环境的有效措施。而果实部分除保护种子外,往往兼有贮藏营养和辅助种子散布的作用。
人们利用各种植物的果实和种子作为食物和提供工业、医药原料等。供食用的果实和种子种类极多,日常生活所不可缺少的主粮、副食,不管是稻米、面粉、瓜果、豆类,极大部分都是植物的种子和果实部分,这里就无需一一举例详述。多种植物的果实和种子是工业上的原料。种子和果实中所贮藏的淀粉、蛋白质、油脂经过提炼后,可用于食品工业和油脂工业,如食用的淀粉、椰油、豆油、菜油,和供饮料的可可、咖啡,以及供工业用的棉籽油、蓖麻油、桐油、乌桕油、乌桕蜡等等。供医药用的果实和种子种类也不少,如蓖麻、巴豆、石榴、木瓜、使君子等。这些将在下册中结合各科、各植物种类详述。
以下就种子和果实的形成、结构和种类,分别加以叙述。
一、种子的形成
种子的结构包括胚、胚乳和种皮三部分,是分别由受精卵(合子)、受精的极核和珠被发育而成。大多数植物的珠心部分,在种子形成过程中,被吸收利用而消失,也有少数种类的珠心继续发育,直到种子成熟,成为种子的外胚乳。虽然不同植物种子的大小、形状;以及内部结构颇有差异,但它们的发育过程,却是大同小异的。
(一)胚的发育
种子里的胚是由卵经过受精后的合子发育来的,合子是胚的第一个细胞。卵细胞受精后,便产生一层纤维素的细胞壁,进入休眠状态。受精卵休眠时期的长短,随植物种类而异,有仅数小时的,如水稻在受精后4—6小时便进入第一次合子分裂;小麦为16—18小时;也有需2—3天的,如棉;有的需延长到几个月的,如茶、秋水仙等。以后,合子经多次分裂,逐步发育为种子的胚。一般情况下,胚发育的开始,较迟于胚乳的发育。
合子是一个高度极性化的细胞,它的第一次分裂,通常是横向的(极少数例外),成为两个细胞,一个靠近珠孔端,称为基细胞;另一个远珠孔的,称为顶端细胞。顶端细胞将成为胚的前身,而基细胞只具营养性,不具胚性,以后成为胚柄。两细胞间有胞间连丝相通。这种细胞的异质性,是由合子的生理极性所决定的。胚在没有出现分化前的阶段,称原胚(proembryo)。由原胚发展为胚的过程,在双子叶植物和单子叶植物间是有差异的。
胚柄在胚的发育过程中并不是一个永久性的结构,随着胚体的发育,胚柄也逐渐被吸收而消失。过去对胚柄的认识只是认为起着把胚伸向胚囊内部合适的位置以利胚在发育中吸收周围的养料,近年来从胚柄细胞的亚显微结构以及不同植物胚柄的有无、长短等方面的差异,有人认为胚柄细胞还能起到另外的一些作用。比较通常的认识是:胚柄除上述作用外,还可能从它的周围吸收营养转运到胚体供其生长发育,以及作为重要养分和调节胚体生长物质的供应源。
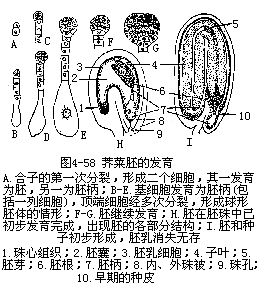
1.双子叶植物胚的发育 双子叶植物胚的发育,可以荠菜为例说明。合子经短暂休眠后、不均等地横向油裂为2个细胞,靠近珠孔端的是基细胞,远离珠孔的是顶端细胞。基细胞略大,经连续横向分裂,形成一列由6—10个细胞组成的胚柄,这些细胞之间有胞间连丝沟通。电子显微镜观察胚柄细胞壁有内突生长,犹如传递细胞,细胞内含有未经分化的质体。顶端细胞先要经过二次纵分裂(第二次的分裂面与第一次的垂直),成为4个细胞,即四分体时期;然后各个细胞再横向分裂一次,成为8个细胞的球状体,即八分体(octant)时期。八分体的各细胞先进行一次平周分裂,再经过各个方向的连续分裂,成为一团组织。以上各个时期都属原胚阶段。以后由于这团组织的顶端两侧分裂生长较快,形成二个突起,迅速发育,成为2片子叶,又在子叶间的凹陷部分逐渐分化出胚芽。与此同时,球形胚体下方的胚柄顶端一个细胞,即胚根原细胞(hypophysis),和球形胚体的基部细胞也不断分裂生长,一起分化为胚根。胚根与子叶间的部分即为胚轴。这一阶段的胚体,在纵切面看,多少呈心脏形。不久,由于细胞的横向分裂,使子叶和胚轴延长,而胚轴和子叶由于空间地位的限制也弯曲成马蹄形(图4-58)。至此,一个完整的胚体已经形成,胚柄也就退化消失。
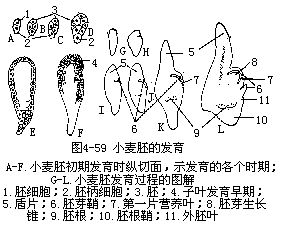
2.单子叶植物胚的发育 单子叶植物胚的发育,可以禾本科的小麦为例说明。小麦胚的发育,与双子叶植物胚的发育情况有共同处,但也有区别。合子的第一次分裂是斜向的。分为2个细胞,接着2个细胞分别各自进行一次斜向的分裂,成为4细胞的原胚。以后,4个细胞又各自不断地从各个方向分裂,增大了胚体的体积。到16—32细胞时期,胚呈现棍棒状,上部膨大,为胚体的前身,下部细长,分化为胚柄,整个胚体周围由一层原表皮层细胞所包围。
不久,在棒状胚体的一侧出现一个小形凹刻,就在凹刻处形成胚体主轴的生长点,凹刻以上的一部分胚体发展为盾片(子叶)。由于这一部分生长较快,所以很快突出在生长点之上。生长点分化后不久,出现了胚芽鞘的原始体,成为一层折叠组织,罩在生长点和第一片真叶原基的外面。与此同时,在胚体的子叶相对的另一侧,形成一个新的突起,并继续长大,成为外胚叶。由于子叶近顶部分细胞的居间生长,所以子叶上部伸长很快,不久成为盾片,包在胚的一侧。
胚芽鞘开始分化出现的时候,就在胚体的下方出现胚根鞘和胚根的原始体,由于胚根与胚根鞘细胞生长的速度不同,所以在胚根周围形成一个裂生性的空腔,随着胚的长大,腔也不断地增大。
至此,小麦的胚体已基本上发育形成。在结构上,它包括一张盾片(子叶),位于胚的内侧,与胚乳相贴近。茎顶的生长点以及第一片真叶原基合成胚芽,外面有胚芽鞘包被。相对于胚芽的一端是胚根,外有胚根鞘包被。在与盾片相对的一面,可以见到外胚叶的突起(图4-59)。有的禾本科植物如玉米的胚,不存在外胚叶。
(二)胚乳的发育
胚乳是被子植物种子贮藏养料的部分,由2个极核受精后发育而成,所以是三核融合(triplefusion)的产物。极核受精后,不经休眠,就在中央细胞发育成胚乳。
胚乳的发育,一般有核型(nucleartype,non-cellulartype)、细胞型(cellulartype)和沼生目型(helobialtype)三种方式。以核型方式最为普遍,而沼生目型比较少见,只出现在沼生目(Helob-iales)植物的胚乳发育中。
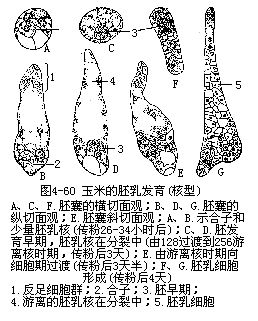
核型胚乳的发育,受精极核的第一次分裂,以及其后一段时期的核分裂,不伴随细胞壁的形成,各个细胞核保留游离状态,分布在同一细胞质中,这一时期称为游离核的形成期(freenuclearformationstage)。游离校的数目常随植物种类而异,多的可达数百以至数千个,才过渡到细胞时期,如胡桃、苹果等。少的仅8或16个核,甚至只有4个核,如咖啡。随着核数的增加,核和原生质逐渐由于中央液泡的出现,而被挤向胚囊的四周,在胚囊的珠孔端和合点端较为密集,而在胚囊的侧方仅分布成一薄层。核的分裂以有丝分裂方式进行为多,也有少数出现无丝分裂,特别是在合点端分布的核。
胚乳核分裂进行到一定阶段,即向细胞时期过渡,这时在游离核之间形成细胞壁,进行细胞质的分隔,即形成胚乳细胞,整个组织称为胚乳。核型胚乳发育的植物种类里,有的是全部游离核都转为胚乳细胞;也有仅胚囊的周围形成一二层细胞,而中央仍保持游离核状态;或是细胞只限于在胚囊的珠孔端形成;仅少数种类是不形成细胞的。单子叶植物和多数双子叶植物属于这一类型(图4-60)。
细胞型胚乳的发育不同于前者的地方,是在核第一次分裂后,随即伴随细胞质的分裂和细胞壁的形成,以后进行的分裂全属细胞分裂,所以胚乳自始至终是细胞的形式,不出现游离核时期,整个胚乳为多细胞结构。大多数合瓣花类植物属于这一类型(图4-61)。
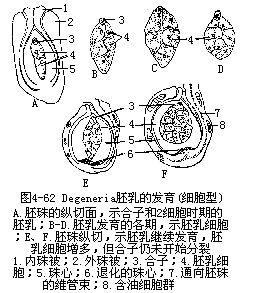
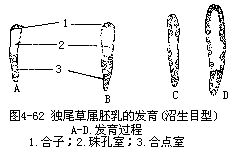
沼生目型胚乳的发育,是核型和细胞型的中间类型。受精极核第一次分裂时,胚囊被分为2室,即珠孔室和合点室。珠孔室比较大,这一部分的核进行多次分裂,成游离状态。合点室核的分裂次数较少,并一直保持游离状态。以后,珠孔室的游离核形成细胞结构,完成胚乳的发育。属于这一胚乳发育类型的植物,仅限于沼生目种类,如刺果泽泻、慈姑、独尾草属(Eremurus sp.)等(图4-62)。
因为胚乳是三核融合的产物,它包括2个极核和1个精子核,含有三倍数的染色体(由母本提供2倍、父本提供1倍),所以,它同样包含着父本和母本植物的遗传性。而且它又是胚体发育过程中的养料,为胚所吸收利用,因此,由胚发育的子代变异性更大,生活力更强,适应性也更广。
种子中胚乳的养料,有的经贮存后,到种子萌发时才为胚所利用的,这类种子有胚乳,称为有胚乳种子。如前面提到过的禾本科植物种子、蓖麻种子等。但另有一些植物,随着胚的形成,养料随即被胚吸收,贮存到子叶里,所以种子成熟时已无胚乳存在,这些是无胚乳种子,如豆类、瓜类的种子。
一般植物种子,在胚和胚乳发育过程中,要吸收胚囊周围珠心组织的养料,所以珠心一般遭到破坏而消失。但少数植物种类里,珠心始终存在,并在种子中发育成类似胚乳的另一种营养贮藏组织,称为外胚乳(prosembryum)。外胚乳具胚乳的作用,但来源与胚乳不同。有外胚乳的种子,可以是无胚乳结构的,如苋属、石竹属、甜菜等;也可以是有胚乳结构的,如胡椒、姜等。
被子植物中的兰科、川苔草科、菱科等植物,种子在发育过程中极核虽也经过受精作用,但受精极核不久退化消失,并不发育为胚乳,所以种子内不存在胚乳结构。