����� �������������ܾ�
һ������
�����������еĻ��ۺʹ����е����Ҵﵽ�����ʱ�ڣ����Ƕ���֮һ�Ѿ����죬��ʱԭ���ɻ���������ס�Ļ��ſ���¶���ơ����Ϊ��һ���Ĵ�����������һ�����Ϊ������anthesis������Щֲ��Ļ������������ſ������Ѿ���ɴ������ã�������һ�������ܾ����ã����ڱջ����۵�ֲ����������Լ�����
��������ֲ��Ŀ���ϰ�Բ�ȫ��ͬ����ӳ��ֲ��Ŀ������䡢�������ںͻ��ڳ��̣��ܲ�һ�¡����磬һ����������ֲ�һ�����������¾��ܿ�����һ���н�����һ�Σ�����������ֲ���ή��л��������ֲ���ڴﵽ�����������ÿ�굽ʱ�����������ꡣ����ֲ��Ŀ������������кܴ���죬��3—5��ģ���������10—12��ģ���������20—25��ģ���������������Ƕ�����ֲ���һ������ֻ����һ�Σ�������ȥ����ֲͬ��Ŀ��������䲻��ȫ��ͬ���������ϼ������紺���ڵĽ϶ࡣһ��˵��������ֲ����Ҷ��������Ҳ����Ҷ�����ģ�����÷�������ȡ��е�ֲ���ڶ��쿪����Ҳ�������Ͽ����ģ������������ڻ��ڵij���Ҳ���в��죬�еĽ����죬���ҡ��ӡ���ȣ�Ҳ�г���һ�������»����������÷���е�һ��ʢ���Ժ�ȫ�����䣬�еij־õ�½�����ţ����ޡ����ѵȡ��ȴ�ֲ������Щ���༸�����꿪������ɿɡ����������ʵȡ�����ֲ��Ŀ���ϰ��������ԭ���ص����������йأ���ֲ�ﳤ����Ӧ�Ľ����Ҳ�����ǵ��Ŵ��������ġ�
��������ʱ�Ļ��䣬һ��ơ������Ѿ����죬����Ļ�����ͨ��һ����ʽ���Ѳ�ɢ�����ۡ������ҵĿ��ѷ�ʽ�Ƕ����ģ�����ͨ�ķ�ʽ�����ѣ������к��ѡ����ѡ����ѵȣ����ڲ�ֲͬ���������м���������ǰ����ɢ�ų����Ļ���ϸ�����������ģ������˵��¶ȡ�ˮʪ�����£�����һ��ʱ�ڵ��ȷ��������¡��ɺ����������ˮ��һ�������ƻ����۵���������ͬʱ����ͷ�ķ������ò���������Ӱ�죬���������ڿ���ʱ�������¡��ɺ�����������ȶ����������ᵼ�¼�����
��������
�����ɻ�����ɢ���ij��컨�ۣ�����һ����ý�������������͵�ͬһ������һ���Ĵ�����ͷ�ϵĹ��̣���Ϊ���ۣ�pollination����
����������������ֳ������ȱ�ٵĻ��ڣ�û�д��ۣ�Ҳ�Ͳ���������ܾ����á���Ϊ������ֳ�����еĴ�����——��ϸ�����Dz�����������ģ��������������ӷ����ڵ������Ҫ���ȫ��������ֳ���̣���һ������ʹ����������——��ϸ���Ļ���������ӽ������۾���������һ�����á�
������Σ����۵��ȷ���Ҫͨ����ͷ�������ʵĴ̼����ú�������������һ������£�����ֻ���䵽��ͷ�ϲ�����Ȼ���ȷ���������ͷ��ɢ��������IJ�ֲͬ�ﻨ�۾���ѡ�����ã�ֻ��һ���������͵Ļ��۲��ܵõ��ȷ����������Ļ��ۻ��ܵ���ͷ�����ƣ���������ֳ���á�����Ϊ������ֳ�����л��۵���Դ���죬�ܾ����������������ͬ���Ľ��Ҳ�Ͳ�ͬ��������һ������������Ӧ�Ե�ǿ����Ҳ�������ʵIJ�ͬ�����ֲ��졣���ֲ����Ŵ��ͱ����������й�ϵ�ġ�
������һ���Ի��������컨���ۼ�������ѧ����
������������һ�������ַ�ʽ��һ���Ի����ۣ�self-pollination������һ���컨���ۣ�cross pollination���������ִ��۷�ʽ����Ȼ�綼�ձ���ڡ�
 ����
����
����1���Ի����� ���۴ӻ�����ɢ�����䵽ͬһ������ͷ�ϵĴ�������Ϊ�Ի����ۡ�����ֲ���еĴ���С���㶹�����ѵȣ������������۵ġ���ʵ��Ӧ���ϣ��Ի����۵ĸ����ָ�ڹ���������ͬƷ�ּ�Ĵ��ۣ�����ũҵ��ͬ���컨��Ĵ��ۡ�
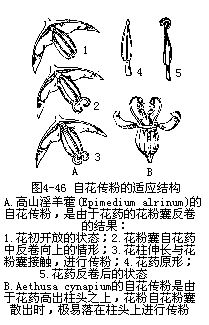
�����Ի�����ֲ��Ļ���Ȼ�ǣ���1�����Ի����������ﳣΧ�ƴ�����������Ұ��úܽ������Ի����������ڱ�������ͷ�ϣ���2������Ļ����Һʹ�������ұ�����ͬʱ����ģ���3���������ͷ���ڱ����Ļ����ȷ��ͻ��۹��������ӵķ���û���κ������谭��ͼ4��46����
�������۷�ʽ�еıջ����ۺͱջ��ܾ���cleistogamy����һ�ֵ��͵��Ի����ۣ�����һ��Ŀ������ۺͿ����ܾ���chasmogamy���Dz�ͬ�ġ�����ֲ��Ļ����������ſ������Ѿ�����ܾ����á����ǵĻ���ֱ���ڻ��������ȷ������۹ܴ��������ҵıڣ�����ͷ����������ܾ�����ˣ��ϸ�ؽ������ڴ�����һ���ڣ������㶹�����ӡ��仨���ȡ��ջ��ܾ�����Ȼ����һ�ֺ�������Ӧ����ֲ���ڻ������������ڿ�������ʱ���ջ��ܾ����ֲ�����һ���㣬�����ֳ���̣����һ��ۿ��Բ�������ˮ����ʪ���������ʳ��
����2.�컨����һ�仨�Ļ��۴��͵�ͬһֲ���ֲͬ����һ�仨����ͷ�ϵĴ��۷�ʽ�����컨���ۡ��ڹ��������ϣ���ͬƷ�ּ�Ĵ��ۺ����������ϲ�ֲͬ���Ĵ��ۣ�Ҳ���컨���ۡ�
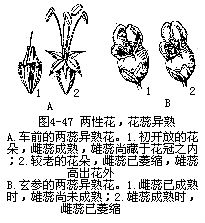
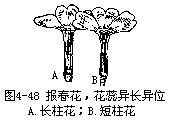
�����컨���۵�ֲ��ͻ����ڽṹ�������ϲ�����һЩ�������Ӧ�Ա仯��ʹ�Ի����۳�Ϊ�����ܣ���Ҫ�����ڣ���1�������ԣ������Ǵ�������ֲ���2�����Ի���������ʹ��ﲻͬʱ���죬�ڴơ��������������У�����������ģ������ס�ݫ�ĵȣ����Ǵ�������ģ���ľ������˵ȣ�ͼ4��47������3���ơ������쳤����λ��Ҳ���ܽ����Ի����ۣ��籨������ͼ4��48������4���������ڱ�����ͷ�ϲ����ȷ���������ȫ�����Դﵽ�ܾ��Ľ�������������顢�ҡ��桢ƻ�������ѵȡ�
 ��
��
�����컨������ֲ���Ƚ��ձ�ش����ţ����Ի�������ȣ���һ�ֽ����ķ�ʽ���������ڵ��Ի����۶�ֲ�����к��ģ���ʹ�������������˥�ˣ�����ũҵ����ʵ�����ѵõ�֤��������С�����Ի����۵�ֲ�������������Ի����ۣ�30—40������˥�˶�ʧȥ�����ֵ��ͬ������������10—15����Ի����ۺ�Ҳ�����ͬ�������컨���ۺ������ܾ��Ͳ����������ǵĺ����������ǿ�������������Ӧ�ԡ�����ľ������ڵĹ۲��о���ָ����“�����Ի����۶�ֲ�ﱾ�����к��ģ����컨���۶�ֲ���������”�����Ľ�����ũҵʵ������ȫһ�µġ�
�����Ի����ۡ������ܾ�֮�����к������컨���ۡ������ܾ�֮�������棬����Ϊ�Ի�����ֲ�����������������ӣ��Ǵ���ͬһ���������£������ӵ��Ŵ���ȱ���ֻ����ã������С�������ںϺ�����ĺ������������Ӧ��С�����컨�������ڴơ����������ڱ˴˲���ȫ��ͬ�����������²����ģ��Ŵ��Ծ߽ϴ���죬�ںϺ�����ĺ����Ҳ���н�ǿ������������Ӧ�ԡ�
������Ȼ�Ի�������һ��ԭʼ�Ĵ�����ʽ���Ժ��Ҳ�������棬������Ȼ��ȴ��������������������Ϊ�Ի����۶�ijЩֲ����˵���������ġ����컨����ȱ������ķ硢���ý����������ʹ���۲��ܽ��е�ʱ���Ի������ֲ�����һȱ�㡣������������ָ��������ֲ����˵���������ܾ���������ֳ���ӣ��ܱȲ���ֳ���ӻ�ֳ���������ú�Щ���ο�����Ȼ��û��һ��ֲ���Ǿ����Ի����۵ģ��������м��ܻ��бȽ��ٵ�һ����ֲ�����ڽ����컨���ۡ����ԣ��������Ի����۵�ֲ�����࣬�����ձ���ڡ�
������������ý���ͳ�ý��
����ֲ������컨���ۣ������������������İ��������ܰѻ��۴���������������ͷ��ȥ�����ͻ��۵�ý���з��������桢���ˮ����Ϊ�ձ���Ƿ�����档���ֲ�ͬ�������۵Ļ�����������һЩ�������Ӧ�Խṹ��ʹ���۵õ���֤��
����1����ý�� ���������ͻ��۵Ĵ��۷�ʽ�Ʒ�ý��anemophily�����������ʽ���۵Ļ����Ʒ�ý����anemophilous flower�����ݹ��ƣ�Լ��1/10�ı���ֲ���Ƿ�ý�ģ��ֺ̱���ֲ���ľ��ֲ���е��ݡ����ľ�ȶ��Ƿ�ýֲ�
������ýֲ��Ļ����ܼ�����״�����|�軨��ȣ��ܲ����������ۣ�ͬʱɢ�ţ�����ͼ 4��28��C��������һ�����ᡢ�������⻬�����ױ��紵�͡��̱���ֲ����С��ˮ���ȵĻ�˿�ر�ϸ������ҩ���ھ��������Ƭ֮�⣬�ܷ����Ĵ�����ʹ�������۴�ɢ��������ȥ����ý���Ļ��������ϳ�����ͷ������״���߳����⣬���ӽ��ܻ��۵Ļ��ᡣ������ýֲ������Ҷ������ϰ�ԣ���������֦�ϵ�Ҷչ��֮ǰ��ɢ���Ļ����ܷ紵��ʱ�����Բ�����֦Ҷ���赲�����⣬��ýֲ��Ҳ���Ǵ����컨�����꣬��������ʧ��������ζ��ɫ����Щ�����DZ�Ҫ���������еķ�ý�����������Եģ�Ҳ��������̱���ֲ��Ļ������Եģ��㡢�ʵ�ֲ��Ļ�Ҳ������
����2����ý�� ������Ϊý����д��۷�ʽ�ijƳ�ý��entomophily�����������ʽ���۵Ļ����Ƴ�ý����entomophilous flower���������л�ֲ�����������洫�۵ģ������Ĵ��������з��ࡢ���ࡢ���ࡢӬ��ȣ���Щ���������ڻ���֮�䣬����Ϊ���ڻ��в��ѣ������Ի���Ϊ��Ϣ���������Dz�ʳ���ۡ�������Ϊʳ�ϡ�����Щ��У����ɱ����Ҫ�뻨�Ӵ�������Ҳ�ͽ����۴��ͳ�ȥ��
������Ӧ���洫�۵Ļ���һ���������������
������ý������������ζ���������档��ֲͬ��ɢ������ζ��ͬ��������������������Ҳ��һ������ϲ����ģ�Ҳ��ϲ����ġ�
������ý������ܲ���֭�����ٻ��Ƿֲ��ڻ��ĸ������֣����Ƿ�չ����������١����۾����ں�����ڻ��ĵײ������еľ��ڡ����۱�¶����ģ������ɼ׳桢Ӭ�Ͷ��ǵķ��ࡢ��������������������ڻ���֮�ڵģ���Ϊ���ǵĵ���Ͷ�������ȡ������ȡ��ʱ��������ճ�����������϶���������ȥ��
������ý������һ�ص��ǻ�������������и�������ɫ�ʡ�һ����俪�ŵĻ���졢�ơ��ϵ���ɫ�������俪�ŵĶി��ɫ��ֻ��ҹ���Ķ�����ʶ�𣬰������ۡ�
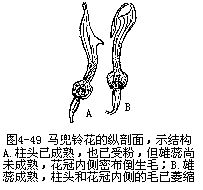
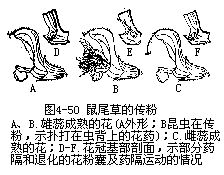
�������⣬��ý���ڽṹ��Ҳ���ʹ��۵�������γɻ�Ϊ��Ӧ�Ĺ�ϵ��������Ĵ�С�����Ρ��ṹ����Ϊ���뻨�Ĵ�С���ṹ�����ٵ�λ�õȣ�����������صġ����������廨������������Ϊ��Ͳ�����ơ��������죬����λ�ڻ�Ͳ���������⣬�ڻ�Ͳ�ڱ�����б������ĵ�ë����Щ��������Ĵ���������ء�������Ĵ����ǿ�һЩС����Ϊý��ģ������ڴ������ʱ��С��˳�ŵ�ë���뻨Ͳ�������ۣ���ʱ����Я���Ļ��۾ͱ����͵��������ͷ�ϡ���Ϊ��Ͳ�ڱڵĵ�ë��δ��ή��С��Ϊ��ë���ڻ��ڣ�һʱ��������ֱ����ҩ���죬����ɢ������ë����ή��Ϊ�����������ͨ�������������������Ҳ��ճ�ϴ������ۣ���������һ������ʱ���ְѻ��۴�����һ������ͷ��ȥ��δ�ڷ�ʱ������Ļ�����ֱ���ģ���������ɺ�����ͼ4��49����������β�ݵĴ��ۣ�Ҳ������Ȥζ�ġ���β�������ο�ֲ����Ļ��ࡢ���ڶ������ɹ�״����5Ƭ������ϲ�ȴ���ѳɴ��Σ���2Ƭ�ϳ�ͷ��״���ϴ�����3Ƭ���ϣ��γ��´�����ˮƽ����������ϴ���������2ö�����1������������ṹ���⣬��Ϊ1����ĸܸ�ϵͳ������ҩ���ӳ��ɸܸ˵ı����ϱ۳���������2������Ļ����ң��±۶̣������Ҳ������չΪ��Ƭ״��2����ı�Ƭ״�±�ͬλ�ڻ��ڹܵĺ�������ס���ڹܵ���ڡ����۷���뻨�ڹܵ������ʱ����Ҫͣ�����´��ϣ�Ȼ����ͷ���ƶ���Ƭ�����ܽ��뻨������ȡ���ۣ����ڸܸ˵ĵ���������Ƭ�����ƶ�ʱ���ϲ��ij�������������ʹ���˵Ļ�ҩ���䵽�۷�ı���������Ҳ��ɢ�������汳�ϡ�����������β�ݵĻ����϶̣������۳���ɢ���Ժ�����ʼ�쳤����ͷ���ôﵽ���汳����λ���ϣ��ȵ����л��۵���һ�۷������һ���ڲ���ʱ�����ϵĻ�������Ϳ�����µ���ͷ�ϣ���ɴ������ã�ͼ4��50����������˵����ý��������Ӧ�ṹ�����ӻ��ܶ࣬�Ͳ�һһ�����ˡ�
������ý���Ļ�����һ��ȷ�ý����Ҫ������ڴֲڣ����д�ͻ����ҩ�ѿ�ʱ��Ϊ�紵ɢ������ճ�ڻ�ҩ�ϣ������ڷû�����ʱ���״������������ܣ��������ͷҲ����ճҺ���ڣ�����һ���Ӵ�������ճס����������ҲԶ�Ϸ�ýΪ�١�
 ��
��
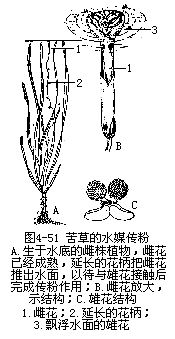
����3���������۷�ʽ ����ý�ͳ�ý�����⣬ˮ������ֲ���еĽ����塢���塢ˮ��ȶ��ǽ�ˮ�������۵ģ����ഫ�۷�ʽ��ˮý��hydrophily������������ֲ���Ǵ�������ģ�����������ˮ�ף����ۻ�����ʱ�������ۻ��Ի������䣬����ˮ�濪�ţ�ͬʱ�ƻ�����Ѹ���ӳ����Ѵƻ�����ˮ�棬���ۻ�Ʈ���ƻ�ʱ�����ֻ���ˮ����������ͷ���ۻ���ҩ�Ӵ�����ɴ��ۺ��ܾ����̣��Ժ�ƻ��Ļ������¾���������״���Ѵ������ˮ�ף���һ�������ɹ�ʵ�����ӣ�ͼ4��51����
��������������ഫ�۵ij���ý��ornithophily�������۵���һЩС�εķ���Heliothrix au-rita����ͷ���г������ȡ����ʱ�ѻ��۴�����ͼ4��52������ţ�������С����Ҳ�ܴ��ۣ�����������
 ��
��
�����������˹������ڷ�
�����컨�������������ܵ��������������ƣ��ò������۵Ļ��ᣬ���ý����û�з磬��ý������������µͣ���ȱ���㹻����ɳ�����۵ȣ��Ӷ����ʹ��ۺ��ܾ��Ļ��ᣬӰ�쵽��ʵ�����ӵIJ�������ũҵ�����ϳ������˹������ڷ۵ķ������Կ˷������������ʹ���۵ò�����֤��ȱ�ݣ��ԴﵽԤ�ڵIJ�������Ʒ�ָ�׳�Ĺ����У�Ҳ��Ҫ��ȡ�˹������ڷۣ��ԴﵽԤ�ڵ�Ŀ�ġ��˹������ڷۿ��Դ���������ͷ�ϵĻ�������ʹ�����������ļ�����������������ӣ�ø�ķ�ӦҲ��Ӧ���˼�ǿ���ٽ������ȷ��ͻ��۹����������ã��ܾ��ʿ��Եõ��ܴ���ߡ���������һ�����������£������������죬���������ʱ�ѵò�����ʱ�Ĵ��ۣ�������붥�������γ�ȱ���������˲������˹������ڷ۾��ܿ˷���һȱ�㣬ʹ�������8—10%���ҡ��������տ�����Ȼ���������£��ձ����϶࣬��������˹������ڷۣ�ͬ������߽�ʵ�ʺͺ�������
�����˹������ڷ۵ľ��巽�����ڲ�ͬ���ﲻ��ȫһ����һ�����ȴ������ϲɼ����ۣ�Ȼ������������ͷ�ϣ����߽��ռ��Ļ��ۣ��ڵ��º���������¼��Ա�أ������Ժ����á�
�����ܾ�
��������������ɺ��۱�����ͷ���ȷ��ɻ��۹ܣ����ڲ�������������——���ӣ�ͨ�����۹ܵ��쳤��ֱ������������ڲ�������ϸ���ͼ��˻����ںϡ������������ӻ����ںϵĹ��̣����ܾ����ã�fertilization������������ֳ���̵���Ҫ�Ρ��ܾ���������һ������Ϊ���ӡ�
������һ������������ͷ�ϵ��ȷ�
����������ͷ�ϵĻ�����������ͷ���ڵ�ճҺ��ճס���Ժ۵��ڱ����ȷ��״�����ͻ�����������쳤���γɻ��۹ܣ���һ���̣��ƻ��������ȷ�����ʹ���������ȷ��������ɻ��۹ܵ������Ƕ��ģ�������ͷ�ķ�����ͻ��۱��������ø�ʹ�л���ͷ���ڵ�ճ�����ʣ����Դ�ʹ�����ȷ�������ֹ�������ڸ����������ճ�����ʵ���Ҫ�ɷ���ˮ�����ࡢ���ܲ��ء�����ø��ά���صȡ����ڷ��������ɳɷ���ֲ��������죬�����������ͷ�ϵĸ���ֲ�ﻨ�۲�����Ӱ��Ҳ�Ͳ�ͬ��
��������������ͷ�Ϻ���������Χ��ȡˮ�֣���ˮ��Ļ�������������Ѹ����ǿ���ۺ������������࣬�����ʵĺϳ�Ҳ����������ߡ�ͬʱ������������ˮ������������߶�����Ҳ��ǿ��Ծ�������ܶ����С�ݣ����Ŷ���ø��������ʣ�ѭ��ϸ������ǰ�����ķ����ͷų�С�ݣ����뻨�۹ܱڵĽ��ɡ���ˮ�Ļ�����Ӫ��ϸ����Һ�ݻ���ǿ��ϸ���ڲ��������࣬ϸ������ѹ���ӣ������ʹ���������ڱ�����һ���������ȷ���ͻ�����γɻ��۹ܣ�pollen tube����ͼ4��53����
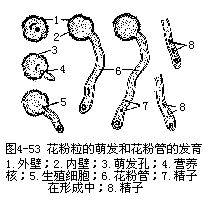
������ֲͬ���ͷ�ķ������ڳɷֺ�Ũ���ϳ�������ͬ���ر��Ƿ������ʵı仯���Ի����ȷ������ٽ������Ƶ�“ѡ��”���á����⣬ʵ��֤����������ӶԻ��۹����������ã�����Լ��ٻ������ѣ�����۵��ȷ��ʣ�����ʹ���۹������������յ����۹ܲ���һ���������ԡ��е�ֲ�����ʱ��ͷ�ϵ�ë״���ͷų����ַ��㻯��������յ������ȷ������á��������������ø���ִ�л�����ǻ������ȷ����������أ��������ڻ��۱ڵĶ���ˮ��ø������ˮ�Ӵ����ɱ����˳����Ի����ȷ��ͻ��۹ܳ�����ͷ������Ҫ���á����绨�����ͻ��۹��еĽ���ø����ʹ��ͷ������ͷ״ͻ��Ľ����ܽ⣬Ϊ���۹ܵ���������ͨ���������ڵĴ�л���ʣ���Ϊ���۹ܵ���������ṩ���ʻ�����
�����䵽��ͷ�ϵĻ�����Ȼ�ܶ࣬������ȫ�����ȷ��ģ��κ�һ��ֲ�↑��ʱ���Խ��ܱ���ֲ��Ļ��ۣ�ͬʱҲ���ܽ��ܲ���ͬ��ֲ��Ļ��ۡ�������ͬ�ֵģ����ڣ����Dz�ͬ�ֵģ��ּ䣩��ֻ�н�������ױ����Ŵ����Ͻ�Ϊ�ӽ�������Ȳ�����Ҳ����С�����п���ʵ�����ԵĽ��䣬�����˵�������ֲ��㷺�ر���Ϊͬһ���ڵ��컨�ܾ��ǿ��͵ģ������Ŵ��ϲ����ش������¾Ͳ����͡����͵Ļ�������ͷ�ϻ��Dz����ȷ��������ȷ��۹��������������ܴ�����ͷ�����ǻ��۹��ڻ����ڵ������ܵ����ƣ����ܴﵽ�ӷ������Դӻ����䵽��ͷ�Ϻ���ͷ�Ի��۾ͽ���“ʶ��”��“ѡ��”�����͵Ļ�������“�Ͽ�”�����͵ľ�����“�ܾ�”��
��������ܾ��ͻ��͵�������ʲô���ݽ�������ϸ������ѧ�ĽǶ��о���֤�������������֯���ʶ��Ӧ�Ǿ����ڻ��۱��Ϻͱ��ڵĵ����ʺ���ͷ���浰���ʱ�Ĥ֮������ϵ���⼸�ֵ�������ʶ���ͻ��͵����ʻ��������컨������Χ�ĵ���������ڵ����ڱڵ������֣�ǰ���ǻ����ڳ���ʱ�ɻ����ұڵ���ձ��ϸ�������ڲ��������ڻ�����ڵ�ǻ϶�ڣ������ǻ����ڳ���ʱ�ɻ���ϸ�����������������ڻ����ڱ��ϵģ����ȷ��������Ϊ�ḻ�����⣬����ͷ���渲��һ����ˮ�ĵ����ʱ�Ĥ�����ճ��Ĥ������ճס������������“ʶ��”���۵�������������������ͷ���ܵ���ʪʱ����ڵ����ڼ������ڱ��ͷų��������ڱڵ���ͨ���ȷ���Ҳ�ӻ����ݳ��������ֻ��۵�����ڻ��۸�������ͷ���棬����ͷ����ĵ��������á��͵Ļ����ܴ���ͷ��ˮ����ʼ�ȷ������͵Ļ����“�ܾ�”����ʱ�ڻ����ȷ����ڲſ�ʼ����Ļ��۹ܶ��γ�������������ֹ�˻��۹ܵ��������е�������ͷ�������ͻϸ���ڲ������������������������۹ܵĴ��롣
�������Խ����ӽ������������ܾ��IJ����ԣ����²��У������ֹ���������ѣ����Խ������ڿ˷������Ե��ϰ�����������о������ж��ִ�ʩ���Բ��ã��磺�û�ϻ����ڷۣ��������ڷۣ��ڷ�ǰ�س���ͷ��ض̻������ӷ����ڷۻ��Թ��ܾ��ȡ�
������������ͷ���������ȷ��ģ������ס��ݵȣ�������Ҫ������������������һЩʱ�����ȷ��ģ�������С����˵ȡ�����ʪ�ȹ��ߣ������¹��ͣ����ܴﵽ�ȷ�����Ҫ��ʪ�Ȼ��¶�ʱ���ȷ��ͻ��ܵ�Ӱ�졣����ʱ���������������������Ž����ڷۣ�ͨ���Dz���ʵ�ġ�������ʪ���漴���Ҳ���������ء����۵�����������ͷ��ά�ֶ�ã������ֹ�����һ���������յ��£������������������⣬�����ֲ����Ŵ���Ҳ�кܴ��ϵ��
�������������۹ܵ�����
����������ͷ�ϵĻ��ۣ��������ͷ�������������͵ģ�������ˮ��ø�Ĵٽ����ú㿪ʼ�ȷ����γɻ��۹ܡ����ڻ�������������ʼ�Ӳ����Χ���ڱ����ܣ�ֻ�����ȷ��ĵط�������չ��أ����Ի��۵�ԭ��������ڱڣ������͵�����£�һ������һ���ȷ���ͻ�����γ�һ��ϸ���Ĺ��ӣ���Ϊ���۹ܡ���Ȼ��Щֲ��Ļ��۾����ȷ��ף�������ơ���«��ֲ��Ļ��ۣ�����ͬʱ�����������۹ܣ���ֻ�����е�һ���ܼ���������ȥ���������;ֹͣ������
�������۹��ж������������ԣ���������ֻ����ǰ��3—5 μm�����γɺ��ܼ����������죬�ȴ�Խ��ͷ��Ȼ���������ӷ���ͬʱ��������ϸ�����ں���ȫ��ע�뻨�۹��ڣ��۹ܶ��˼��У��������ϸ���͵Ļ�������Ӫ���˺�2������ȫ�����뻨�۹��У�����ϸ���͵Ļ�������Ӫ���˺���ֳϸ�����뻨�۹ܺ���ֳϸ�����ڻ��۹��ڷ��ѣ��γ�2�����ӡ�
�������۹�ͨ�����������ӷ�������;�����ɷ�Ϊ���ֲ�ͬ�������һЩֲ��Ļ����м�ɿ��ĵĻ����������۹�������ʱ���Ż������������죬�����ӷ�����һ������ǻ���������������Ϊ�����������֯��transmitting tissue����һ�㱡��ϸ�������������۹�����ʱ�辭��ø�����ã���������֯����֯ϸ�����в�������ܽ⣬���۹ܾ���ϸ��֮��ͨ�������۹��ڻ����е������������û��۱��������������Ӫ���⣬Ҳ�ӻ�����֯��ȡ���ϣ���Ϊ�����ͽ��ɹܱںϳ�����֮�á����۹ܵ����������ڼ�˲��֣��뻨�۹ܶ���ԽԶ�IJ���Խ��˥�ϡ�
���������ȷ��ͻ��۹ܵ������ٶ��ڲ�ֲͬ����������������ı仯���Dz���ȫһ�µģ�����Ӵ��۵��ܾ���ʱ��Ҳ���ϴ�ľ��ֲ��Ļ��۹���������������ƻ���ɴ��۵��ܾ���ʱ��Ϊ5�죬����ֲ�����賤��һ���һ��ࣻ��һ��ũ����Ļ����ȷ����������ٶȽϿ죬����ˮ��ֻ��1.5Сʱ��С��1Сʱ��������15—32Сʱ����Щ������Ҫ���Ŵ�����������������֮�⣬�������ص�Ӱ��Ҳ��ʹ�ٶ������ı䣬�绨�������ĺû�������ʱ���µĸߵͺͿ��������ʪ�ȵȡ�
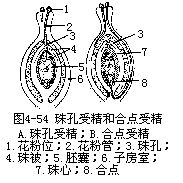
�������۹ܵ����ӷ��Ժ���ֱ��������ף��������ң�ֱ�����飩�����߾��������������������ڣ��������������飩��������������ң�ͳ��Ϊ����ܾ���porogamy��ͼ4��54��A����Ҳ�л��۹ܾ���������ĺϵ�������ҵģ���Ϊ�ϵ��ܾ���chalazogamv��ͼ4��54��B����ǰ����һ��ֲ�����У��������ټ��������ܡ����ҵ��ܾ�������һ���͡����⣬Ҳ�д����鱻���ɲ���������ҵģ����в��ܾ���mesogamy����������ټ������Ϲϡ����ۻ��۹���������ȡ����һ��;�����������ȷ��������������ң���һ����IJ���ԭ��һ����Ϊ�ڴ���ijЩ��֯�����������������������֯��̥�����ӷ��ڱں���ϸ���ȴ���ij�ֻ�ѧ���ʣ����յ����۹ܵĶ���������
��������������ֲ���˫�ܾ����̼�������ѧ����
�������۹ܾ��������������ӷ���ֱ�����飬Ȼ����ף������������ҡ���������֯�ϱ�����������۹ܿ��������������ң��������ĽϺ����������۹�����ͨ����ʵ��������֯�����ܽ������ҡ�
�������۹ܽ������ҵ�;���ڲ�ֲͬ���Dz�һ���ģ���������ϸ����һ����ϵ���д��Ѻ���ϸ��֮��������ҵģ��������д���1����ϸ���У�Ȼ��������ҵģ����ޣ������ƻ�1����ϸ����Ϊ�������ҵ�ͨ·�ģ������ÿ������Ǵӽ������ϸ������ģ������ס����۹ܽ������Һܵ�ĩ�˼������ѣ������Ӽ�����������ע�����ҡ�����ԭ������Ϊ�����������ڵĵ����������£�����ϸ�����Ʋ�Ϊ�Ի��۹���������ֱ�ӵ����ã������۹�����ϸ����ϸ���ʽӴ�ʱ������ѹ����ͻȻ�ı䣬���¹ܵ�ĩ�����ѣ�Ҳ����Ϊ���۹ܹܱڵ��ܽ⣬�緬�ѡ����飬Ҳ��ԭ��֮һ��
 ��
��
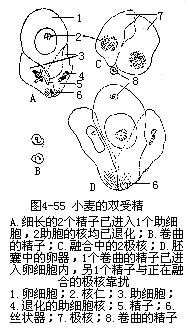
�������۹��е����������ͷŵ������к��ŷ������Ӻ���ϸ���Լ����Ӻ�2���˵��ںϡ�2�����е�1�������ںϣ��γ��ܾ��ѣ���ƺ��ӣ�����������Ϊ�ߡ���1�����Ӻ�2�����ˣ�������ˣ��ںϣ��γɳ�������ˣ��Ժ���Ϊ���顣��ϸ���ͼ���ͬʱ��2�����ӷֱ�����ںϵĹ��̣��DZ���ֲ��������ֳ����������Ϊ˫�ܾ���doublefertilization����ͼ4��55����
�����������ҵ�2�������Ƿ��ڽṹ���������Եȷ�����ȫ��ͬ��ͨ�����������Ĺ۲��о�֤����2���Ӽ����пɲ���IJ�����ڵġ�Russell��1980—1983�������ڵ����������о�������plumbagozeylanica������ʱ���۲쵽2�������е�1����ͻ������Ӫ��ϸ���ĺ��ϣ���1����ͨ��������˿���һ�������������⣬������Ӫ�����ϵ��Ǹ����Ӻ��϶������������壬����1�������϶�����壬��ˣ�������Ϊ�������ӿ����Ƕ��εģ�dimorphic����
��������ϸ����ϵľ��ӣ��ڽ�����ϸ�����Ѻ˽ӽ�ʱ�����˵�Ⱦɫ���������Ѻ˵ĺ�Ĥ�ϣ�Ȼ����ѷ�ɢ��ͬʱ����1��С�ĺ��ʣ��������˺��Ѻ˵�Ⱦɫ���������һ���ۺ˵ĺ���Ҳ�ʹƺ˵ĺ����ں���һ�𣬽�����һ�ܾ����̡���1�����Ӻͼ��˵��ںϹ��������������ӵ��ں��ǻ������Ƶģ����ӳ�ʱҲ�ʾ����Ĵ�״���Ժ��ɿ��뼫�˱���Ӵ���2��Ⱦɫ�ʺ�2���ʺϲ�������������̡����Ӻ��ѵĽ�ϱȾ��Ӻͼ��˽�ϻ��������Ծ��Ӻʹ����˵ĺϲ���ɵý��硣
�����ܾ�ʱ������ϸ����ϸ�����Ƿ������ϸ���У��Ӳ�ͬ�о�����ó��Ľ����Dz�ȫ��ͬ�ġ�һ����Ϊ����ϸ����Ҳ�����ܾ�����һ�ֳ��෴�۵㣬���������������о���֤����һ������Ϊ��ȷ�����������ܾ�ʱ�������ṹ��ʾֻ�����Եĺ˻����ںϣ�������ϸ������������ƻ�����ϸ���С�
�����ܾ��������еķ���ϸ����������Ѷ��������࣬��Ϊ�ߺ����鷢��ʱ�����ϣ������ȫ����ʧ��
��������ֲ���˫�ܾ���ʹ2��������Ĵơ��������ں���һ�𣬳�Ϊ1��������ĺ��ӣ��ָ���ֲ��ԭ�е�Ⱦɫ����Ŀ����Σ�˫�ܾ��ڴ����ױ��Ŵ��ԣ���ǿ������������������Ӧ�Է����Ǿ��нϴ������ġ��������ںϾͰѸ���ĸ�����в�����Ŵ����������һ���γɾ�˫���Ŵ��Եĺ��ӡ��������Ӽ���ͬ�����γɵĺ�������п����γ�һЩ�µı��졣���ܾ��ļ��˷�չ�ɵ�������������ģ�ͬ�����и���ĸ�����Ŵ����ԣ���Ϊ����һ�����ڵ����ϣ�����Ϊ���̺ͷ�չ��һ�ص��ṩ�������������ԣ�˫�ܾ���ֲ�����������ֳ�������������������ʽ��
�������ģ��ܾ���ѡ������
������ͷ�Ի��������ȷ����Լ����ҶԾ���ϸ���Ľ��룬������ѡ��������Ҳ����˵��ֻ���ܺ���ͷ������������������Э���Ļ������������ȷ�����ϸ��Ҳֻ�ܺ���������������Ӧ�ľ����ں���һ�����ԣ�����ֲ����ܾ���������ѡ���Եģ����ֶԻ��ۺ;��ӵ�ѡ���ԣ���ֲ���ڳ��ڵ���Ȼѡ�������±��������ģ�Ҳ�DZ���ֲ����������е�һ����Ҫ������ˣ���Ȼ������ͷ�Ͽ������в�ֲͬ��Ͳ�ֲͬ������Ļ��ۣ����ǣ�ֻ���ʺ�����һ�ܾ����̵�ֲ�ﻨ�ۣ����ܲ���Ч����
�����ܾ����õ�ѡ������Ϊ�������ע�⣬���������ָ�����ܾ��������û��ѡ���ԣ��Ͳ����ܱ��������ܾ��ͽ����ܾ��ĺ�����Ҳ�����ܵõ������ܾ����洦��ʵ��֤����������ò�ֲͬ�꣬������ͬ����Ļ�ϻ��۽����ڷۣ�ֻ�����ʺ�����ͷ�����ҵĻ����о����ȷ��Ŀ��ܣ������˽����Լ����ϵĻ���������ˣ����û���ڷۡ��˹������ڷ�����߲������˷��Խ���ԶԵ�ӽ��IJ����ԣ��Լ���ߺ���Ի�������Ӧ���������ܵ��ձ����ӡ�ѡ���ܾ�������Ϊũҵ����ʵ���������ĺô���ϣ����Ϊѡ�ֺ����ַ��������춨�˻�����
�����ڱ���ֲ���У�˫�����ѺͶྫ���ѵ��������Σ�Ҳ�з��֣����Ӿ��ӽ�����ϸ���ı�����ϸ����ͬ�����ã�ʹ�ߵ�Ӫ���������Ӵ����Ŵ��Է����仯��
�������壩���ں���ֳ����������
��������������£�����ֲ���������ֳ�Ǿ�����ϸ���;��ӵ��ںϣ��Է������ߣ�������Щֲ��������������ںϣ�Ҳ��ֱ�ӷ������ߣ�������������ں���ֳ��apomixis�������ں���ֳ��������ϸ���������ܾ���ֱ�ӷ������ߣ����ѹ�Ӣ������̵ȣ���������ƹ´���ֳ��part-henogenesis������������ϸ��������ϸ���˵ȷ���ֳ��ϸ���������ߣ���С��β�����߲ݵȣ�����������Ϊ��������ֳ��apogamy����Ҳ�е��������Ļ��鱻ϸ��ֱ�ӷ������ߵģ����������Citrus������Ϊ��������ֳ��apospory����
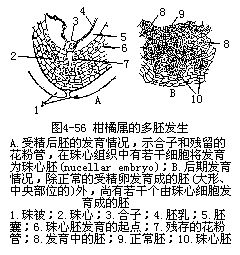
������һ������£�����ֲ�������ֻ����1�����ң�ÿ������Ҳֻ��1����ϸ���������ܾ���ֻ�ܷ�����1���ߡ����е�ֲ��������������2���������ߴ��ڣ���һ�����Ϊ��������polyembryony������������IJ�����������������������ֳ����������ֳ�Ľ����ͼ4��56����Ҳ��������1���ܾ��ѷ��ѳɼ����ߣ�����1�������з���������ҵ�Ե�ʡ�