第四节 胚珠的发育和胚囊的形成
被子植物的有性生殖,除了了解花药和花粉的形成和发育外,还需要了解胚珠和胚囊的形成和发育过程。胚珠是由心皮内表面沿腹缝处形成的突起发展演变而成的,在结构上,一个成熟的胚珠是由珠心、珠被、珠孔(micropyle)、珠柄和合点(chalaza)等几部分组成。在性质上,心皮相当于大孢子叶,胚珠相当于产生大孢子的大孢子囊,就如雄蕊的花药是产生小孢子(花粉粒)的小孢子囊一样。成熟胚珠的珠心内,将产生一个单相核的胚囊细胞,也就是大孢子,以后经过细胞分裂,形成8核、7细胞结构的成熟胚囊(一般情况)。卵细胞就在胚囊里产生。
一、胚珠的发育
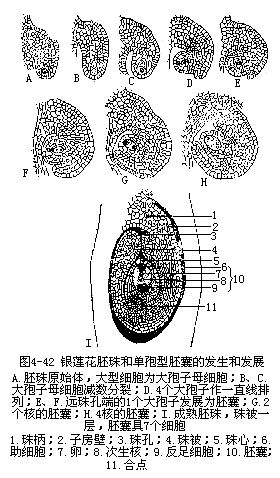
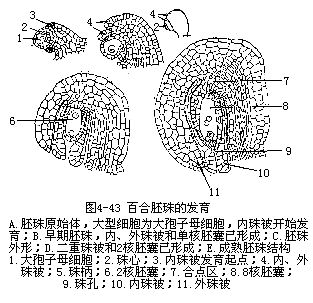
胚珠是在心皮内侧沿腹缝处的胎座上发展起来的,最初产生的一团突起是胚珠的珠心,这是胚珠中最重要的部分,以后的胚囊就是由这部分的细胞发育出来的。由于珠心基部外围的细胞加速分裂,新细胞逐步在珠心周围向上扩展,终于将珠心围在中央,仅在顶端留下一个小孔可以通向珠心。包在珠心周围的细胞层发展为珠被,顶端的小孔称为珠孔。有些植物的胚珠、珠被只有一层(图4-42),如胡桃、向日葵、银莲花属等,但较多植物的胚珠,珠心周围有两层珠被包围,内层为内珠被(inner integument),外层为外珠被(outer integument),外珠被的形成是在内珠被形成之后,按同样方式由胚珠基部发展而成的(图4-43),如小麦、水稻、油菜、棉、百合等。胚珠基部的一部分细胞发展成为柄状结构,与心皮直接相连,称珠柄。胚珠基部与珠孔相对的部位,珠被、珠心和珠柄相愈合的部分称合点。心皮的维管束分支由珠柄进入胚珠,最后到达合点。
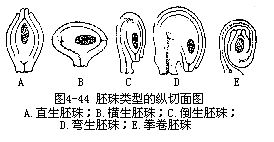
胚珠在生长时,珠柄和其他各部分的生长速度,并不是一样均匀的,因此,胚珠在珠柄上的着生方位,也就有着不同的类型。一种是胚珠各部分能平均生长,胚珠正直地着生在珠柄上,因而珠柄、珠心和珠孔的位置列于同一直线上,珠孔在珠柄相对的一端,这类胚珠,称为直生胚珠(orthotropous ovule),如大黄、酸膜、荞麦等的胚珠(图4-44,A);另一种类型,也是最为习见的类型,是倒生胚珠(anatropous ovule),这类胚珠的珠柄细长,整个胚珠作180°扭转,呈倒悬状。珠心并不弯曲,珠孔的位置在珠柄基部一侧,靠近珠柄的外珠被常与珠柄贴合,形成一条向外突出的隆起,称为珠脊(raphe)(图 4-44,C),大多数被子植物的胚珠属于这一类型。直生胚珠和倒生胚珠可认为是两种基本类型,二者之间尚有一些过渡的型式,如有的胚珠在形成时胚珠的一侧增长较快,使胚珠在珠柄上成90°的扭曲,胚珠和珠柄的地位成为直角,珠孔偏向一侧(图4-44,B)。这类胚珠称为横生胚珠(amphitropousovule)。也有胚珠下部保持直立,而上部扭转,使胚珠上半部弯曲,珠孔朝下,向着基部,但珠柄并不弯曲,称弯生胚珠(campylot-ropousovule)(图4-44,D),如云苔、苋、豌豆、蚕豆和禾本科植物的胚珠。如果珠柄特别长,并且卷曲,包住胚珠,这样的胚珠称为拳卷胚珠(circinotropousovule)(图4-44,E),如仙人掌属、漆树等。
二、胚囊的发育和结构
(一)大孢子的发生
胚珠的珠心原是由薄壁组织的细胞所组成,以后在位于珠孔端内方的珠心表皮下,出现一个体积较大,原生质浓厚,具大细胞核的孢原细胞;在一些植物种类里,孢原细胞可以直接起到大孢子母细胞(即胚囊母细胞)的作用(图4-42,A;图4-43,A)。但有些植物的孢原细胞须经过一次平周分裂,形成一个周缘细胞和一个造孢细胞,前者可以不再分裂,或经平周和垂周分裂后,形成多层珠心细胞,而后者通常不经分裂,直接成为大孢子母细胞。也有孢原细胞不止一个的,但一般仍然只有一个是能够继续发育,成为大孢子母细胞。
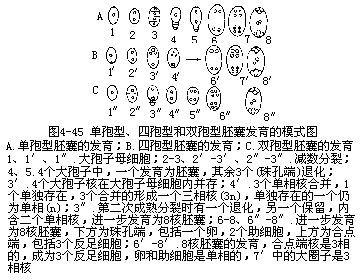
由大孢子母细胞进一步发育为大孢子,可以区别为三种不同的情况:第一种情况是大孢子母细胞经过两次连续的分裂,其中一次是染色体的减数,分裂后,每个细胞产生各自的细胞壁,成为4个含单相核的大孢子。大孢子一般作一直线或T形在珠孔端排列(图4-42,4-45,A)。其中位于珠心深处的1个经过进一步发育,以后成为8核、7细胞的胚囊,其余3个以后退化消失(图4-45,A)如在蓼科植物中所见;第二种情况是大孢子母细胞在减数分裂时的第一次分裂出现细胞壁,成为二分体;二分体中只有一个取得进一步的发育,进入第二次分裂,形成二个单倍体的核,而另一个二分体即退化,以后消失。二分体中保留下来的一个细胞在第二次分裂时并没有形成新壁,所以二个单相核(大孢子核)同时存在于一个细胞中,以后共同参与胚囊的形成(图4-45,C),如葱、慈姑等植物的大孢子发生是经由这一途径的;第三种情况是大孢子母细胞在减数分裂时二次分裂都没有形成细胞的壁,所以4个单倍体的核共同存在于原来大孢子母细胞的细胞质中,以后这4个大孢子核一起参与胚囊的形成(图4-45,B),如贝母、百合等植物的大孢子发生就属这一类型。以后由上述三种大孢子发育成的胚囊分别称为单孢型胚囊、双孢型胚囊和四孢型胚囊。
(二)胚囊的形成
如上所述,由于大孢子起源方式的相异,造成被子植物胚囊形成有不同的类型。但是,被子植物中70%以上的植物胚囊的发育类型是单孢型的,即由一个单相核的大孢子经三次有丝分裂,进一步发育成一个具8个核、7个细胞的胚囊。这种胚囊类型也称蓼型或正常型。现将发育过程概述如下:
直列形的4个大孢子形成后,其中合点端的一个吸收周围营养逐渐长大,而另外3个逐渐退化消失。长大的大孢子也可称单核胚囊,含有大的液泡,最后长大到占有珠心的大部分体积。
当大孢子长大到相当程度的时候,这一单相核即行分裂三次,第一次分裂生成的2新核,依相反方向向胚囊两端移动,以后每个核又相继进行二次分裂,各形成4个核,这三次分裂都属有丝分裂,但每次分裂之后,并不伴随着细胞质的分裂和新壁的产生,所以出现一个游离核时期。以后每一端的4核中,各有1核向中央部分移动,这2个核称为极核(polar nuclei),同时在胚囊两端的其余3核,也各发生变化。靠近珠孔端的3个核,每个核的外面由一团细胞质和一层薄的细胞壁包住,成为3个细胞,其中1个较大,离珠孔较远,称为卵细胞,另2个较小,称为助细胞(synergid),这3个细胞组成卵器(egg apparatus)。另3个位于远珠孔端的细胞核,同样分别组成3个细胞,聚合一起,成为3个反足细胞(antipodal cell)(图4-42;图4-43;图4-45,A)中央的2个极核组成1个大型的中央细胞(central cell)。至此,一个成熟的胚囊出现了7个细胞,即1个卵细胞,2个助细胞,3个反足细胞和1个中央细胞。在有的植物种类里,中央细胞的2个极核在受精前,仍保持分离状态,但也有的植物种类,2极核在受精前就融合为一,例如在棉花中可以见到这一情况。
百合胚囊的结构是植物学教学及实验中经常提及的典型材料,在成熟的胚囊中和蓼型胚囊一样具8个核,7个细胞,但它的发育过程属于四孢型,又称贝母型。具体过程是大孢子母细胞减数分裂后形成4个单倍体的核共同存在于大孢子母细胞的细胞质中,并且呈1和3的排列,即1个核在珠孔端,另3个核在合点端。然后,珠孔端的核进行一次正常的有丝分裂形成2个单倍体的子核,而合点端的3个核分裂时纺锤体相互融合成一个共同的纺锤体,分裂结果形成2个三倍体的子核,它的体积较单倍体的子核大,核仁也多,形状也不同。然后,所有的核各分裂一次,成为8个核。4个在合点端的是三相核,4个在珠孔端的为单相核。接着两端各有一核移向中央。与蓼型胚囊相似,在胚囊的珠孔端发育为1个卵和2个助细胞,在合点端为3个反足细胞,在中部的2个极核构成1个中央细胞。但不同的是它的反足细胞和一个极核是三相的。
在7细胞组成的胚囊(也就是被子植物的配子体)中,卵细胞是最为重要的,它是雌配子,是有性生殖的直接参与者,经受精后的卵细胞将发育成胚。卵细胞有高度的极性,细胞中含有一个大液泡,位于细胞近珠孔的一边;核大型,在细胞中的位置处于液泡的相反一边。卵细胞周围是否有壁包围,不同植物种类的情况是不一样的,玉米和棉花卵细胞的细胞壁仅分布在细胞的近珠孔端部分,其余部分则为质膜所包围,不存在细胞壁;但在荠菜中,整个卵细胞的四周几乎全部为细胞壁所包围,只是在合点端出现细胞壁的不连续状态。卵细胞和两侧的助细胞间有胞间连丝相通。
胚囊细胞中结构最为复杂,并在受精过程中起到极其重要作用的是2个助细胞。助细胞紧靠卵细胞,与卵细胞成三角形排列,它们也有高度的极性,外有不完全的壁包围,壁的厚度同样是不均匀的,近珠孔一边较厚,相反的一端只有质膜包住。细胞内的大液泡位于靠合点的一边,而核在近珠孔端,与卵细胞的情况正相反。助细胞最突出的一点,是在近珠孔端的细胞壁上出现有丝状器(filiform apparatus)结构(参阅图4-55)。丝状器是一些伸向细胞中间的不规则片状或指状突起,这些突起是通过细胞壁的内向生长而形成的,它们的作用使助细胞犹如传递细胞。据多方研究,认为助细胞在受精过程中能分泌某些物质诱导花粉管进入胚囊,同时还可能分泌某些酶物质,使进入胚囊的花粉管末端溶解,促进精子和其他内含物质注入胚囊。此外,从棉花助细胞的亚显微结构的研究证明,助细胞还能吸收、贮藏和转运珠心组织的物质进入胚囊。助细胞的寿命通常较短,一般在受精作用完成后就破坏。
胚囊中反足细胞的数目和形状,以及细胞内核的数目都有很大变化。反足细胞的数目,可以从无到10余个,即使蓼型胚囊的反足细胞,也不一定是3个,常常可以继续分裂成一群细胞。竹亚科的一些种类里,反足细胞可达300个之多。前面提到的贝母型胚囊,反足细胞具三倍体(3n)。反足细胞的核可以是1个,也有具2核或更多核的。反足细胞的寿命通常短暂,往往胚囊成熟时,即消失或仅留残迹,但也有存在时间较长的,如禾本科植物。反足细胞的功能是将母体的营养物质运转到胚囊。
除卵器和反足细胞外,在胚囊里还有1个大的中央细胞,它含有2个极核的内容。正常类型胚囊的极核是2个,但也有1个、4个或8个的。融合的极核也称次生核。绝大部分的中央细胞为 1个大液泡所占据,次生核常近卵器,它的周围有细胞质围绕。2极核的融合,可以发生在受精作用之前,或是在受精过程中。中央细胞与第二个精子融合后,发育成为胚乳。